|
|
 |
|
|
|
|
|
Élément principal constituant le cerveau, les neurones se comptent en centaine de milliard dans
un cerveau adulte normal. Ils sont le sièges de traitement de plusieurs informations parvenant des organes ou du cerveau.
Mis à part leur forme étoilée, l'autre particularité des neurones est le fait qu'ils ne peuvent pas se diviser comme les autres
cellules de notre corps. Tous les neurones entrent en contact les uns avec les autres, et peut-être même avec d'autres cellules
telles que les cellules musculaires.
II-1/
Anatomie et constitution d'un neurone :
Tous les neurones qui forment le
système nerveux ont la même apparence. Chacun d'eux est constitué d'un corps cellulaire où est situé le noyau contenant l'information
génétique. Il a été bloqué autour de ce noyau, il y a des prolongements cytoplasmiques particuliers. On en distingue deux
types. Les uns sont appelés dendrites et sont sous formes de branches d'arbre. Elles possèdent des milliers d'épines qui bourgeonnent
à leur surface, pour faciliter la communication avec les autres neurones. L'autre type de prolongement cytoplasmique est appelé
axone. C'est un unique prolongement qui relie le corps cellulaire aux dendrites d'autres neurones ou aux cellules musculaires
auxquelles est relié le neurone. Sa longueur varie selon l'emplacement du neurone et l'emplacement de la cellule avec qui
il est en contact. Les neurones possédant les axones les plus longs de l'organisme sont ceux qui, situés dans la moelle épinière,
sont en contact avec les pieds. Il se termine par des ramifications appelées terminaisons axonales. Cependant, lorsque l'axone
entre en contact avec une dendrite ou une cellule effectrice (cellule musculaire, glande...), cela ne se fait pas directement,
mais il y a présence d'un espace entre les deux appelé synapse. Cette synapse est celle qui facilite la communication axone/dendrites.
Autour des axones, s'entourent des fois des cellules gliales sous forme de gaine isolante composée essentiellement de myéline.
Cette gaine faite de substance grasse, est appelée gaine de myéline. Elle aide à accélérer la transmission de l'influx nerveux
(ou message nerveux), qui constitue un ensemble d'informations transportées par les fibres nerveuses depuis des récepteurs,
sensibles à des stimuli provenant de l'environnement, jusqu'aux membre effecteurs.
| Un neurone et ses composantes |
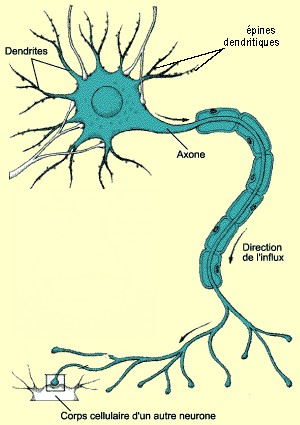
|
| (source: le cerveau à tous les niveaux) |
II-2/ Communication et conduction nerveuse :
Penser, bouger, parler, rire, pleurer,
peur, faim...tout cela est assuré par la communication qui existe entre différents neurones ou entre neurones et cellules
musculaires. Cette communication est soit une conduction électrique, soit une transmission chimique. La conduction électrique
est ce qui permet de faire "voyager" l'influx nerveux à l'intérieur d'un seul neurone, alors que la transmission chimique
est ce qui permet à l'influx nerveux de passer d'un neurone à un autre ou d'un neurone à une cellule musculaire.
| Conduction électrique et transmission chimique (1) |
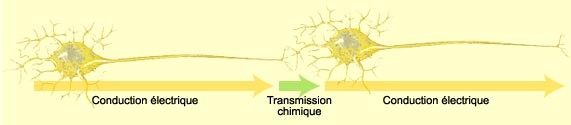
|
| (source: "Le cerveau à tous les niveaux") |
| Conduction électrique et transmission chimique (2) |
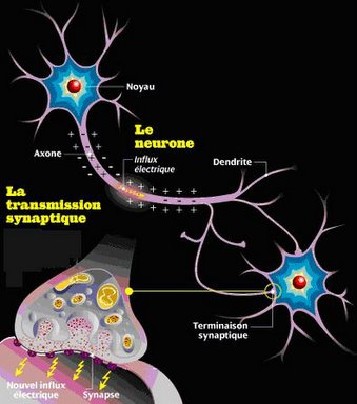
|
| (source: http://www.lexpress.fr/info/sciences/dossier/neurodon/dossier.asp?ida=426750) |
II-2-1/ La
conduction électrique :
La conduction électrique est caractérisée,
comme son nom l'indique, par une activité électrique qui permet la conduction de l'influx nerveux le long du neurone. Ce neurone
obéit à la loi du tout ou rien, c'est à dire qu'il y a un seuil minimum que le stimuli doit atteindre pour qu'il y ait naissance
d'un message nerveux, s'il ne l'atteint pas, on remarque qu'au message au neurone suivant, si ce seuil est atteint, l'activité
neuronale est maximale, quelque que soit l'intensité du stimuli (tant qu'il reste au-dessus du seuil) l'activité neuronale
reste la même. On distingue alors deux types de potentiels: le potentiel de repos, lorsqu'il n'y a pas de conduction de message
nerveux, et le potentiel d'action, lorsqu'il y a conduction d'un message nerveux.
II-2-1-1/ Le potentiel de repos :
La particularité de la membrane
neuronique est sa semi-perméabilité. En d'autres termes, cette membrane a la capacité de laisser passer certains élément chargés,
appelés ions, plus facilement que d'autres.
On appelle potentiel de repos le
moment où il règne un certain équilibre entre les ions à l'intérieur et à l'extérieur de la membrane neuronique. Durant cette
période, les ions qui passent le plus facilement sont les ions K+ (potassium) chargés positivement. Les ions CL- (chlorure)
et Na+ (sodium) ainsi que d'autres grosses molécules chargées négativement circulent plus difficilement.
Des chercheurs ont vérifiés à l'aide
d ’une micro pipette et d’une électrode, le potentiel électrique (mesuré en millivolts ) à l’intérieur du
neurone, et le potentiel électrique à l’extérieur du neurone, lorsqu’il est au repos. Ils ont mesuré une différence
de potentiel électrique entre l’intérieur et l’extérieure d'environ -70 mV. La partie interne du neurone est donc
chargée négativement par rapport à la partie externe. Ceci est dû à la répartition des ions de part et d'autre de la membrane.
L'enregistrement du potentiel de
repos à l'ossilloscope donne le résultat suivant:
| Le potentiel de repos |

|
| (source: schéma fait par nous-même) |
Cet équilibre qui règne au repos
peut être perturbé par l'arrivée d'un stimuli provenant d'un milieu extérieur. Cette perturbation d'équilibre entraîne la
création d'un nouveau type de potentiel, le potentiel d'action.
II-2-1-2/ Le potentiel d'action :
La modification brutale et localisée
de l'équilibre électrique qui régnait de part et d'autre de la membrane neuronique, due à la perception d'un stimuli, est
appelée potentiel d'action. Il s'agit d'une inversion de la polarisation membranaire qui se propage tout au long de l'axone.
C'est-à-dire que l'intérieur, chargé négativement par rapport à l'extérieur, devient chargé positivement. c'est la dépolarisation.
Juste après le passage de l'influx, il y a une repolarisation, c'est-à-dire que la membrane retourne à son statut d'équilibre
électrique caractéristique du potentiel de repos.
Une des particularités du potentiel
d'action est son seuil de déclenchement, que l'on a évoqué plus haut. En effet, la transmission du message neuronal obéissant
à la loi du tout ou rien, il faut un seuil de déclenchement à partir duquel l'influx nerveux parviendra au neurone suivant
de façon optimale (ce qui prouverait le fait qu'il y ait une conduction de l'influx tout le long du premier neurone qui se
fait de façon optimale). Ce seuil est atteint lorsque le potentiel membranaire atteint -3OmV (à partir de -30mV, et quel que
soit l'intensité du stimuli, l'intensité de l'influx nerveux est maximale et ne varie pas).
L'enregistrement du potentiel d'action
à l'ossilloscope donne le résultat suivant:
| Le potentiel d'action |

|
| (source: schéma fait par nous-même) |
Mais cette conduction électrique ne servirait à rien s'il n'y avait pas un autre mécanisme complémentaire
qui permettrait le passage de l'influx d'un neurone à un autre ou d'un neurone à une cellule effectrice. En effet, la communication
entre neurone et cellule réceptrice se fait grâce à la transmission chimique.
II-2-2/ La
transmission chimique :
Comme les neurones ne se touchent
pas au niveau de leurs synapses, il doit y avoir certains organismes qui permettraient le passage du message nerveux de la
cellule pré-synaptique (cellule d'où vient l'influx nerveux) vers la cellule post-synaptique (cellule qui reçoit l'influx
nerveux). Il s'agit alors d'une molécule chimique appelée "neurotransmetteur".
On peut distinguer la cellule pré-synaptique de celle post-synaptique par plusieurs caractéristiques spécifiques. On
remarque que les cellules pré-synaptiques se terminent par un bouton terminal de l'axone au niveau duquel on remarque la présence
de mitochondries et de microtubules qui permettent le transport des neurotransmetteurs du corps cellulaire (où ils sont produits)
jusqu'à l'extrémité de l'axone. Arrivés au bout de l'axone, les neurotransmetteurs se regroupent dans des organites sphériques
appelés vésicules.
Ce sont ces vésicules qui vont permettre
le passage des neurotransmetteurs d'une cellule à l'autre. Ce passage est assuré par un mécanisme appelé exocytose. Il consiste
en la fusion des membranes des vésicules avec celle du bouton pré-synaptique. Ceci permet alors la création de poche desquelles
les neurotransmetteurs s'échappent, pour se retrouver dans la fente synaptique séparant les deux cellules. Celle-ci n'est
pas très épaisse, elle ne mesure que 0,02 microns.
Au niveau du neurone post-synaptique,
on remarque des sortes de récepteurs localisés sur la membrane neuronale. Les neurotransmetteurs, libérés dans la fente synaptique,
se fixent sur ces récepteurs (chaque récepteur est spécifique à un seul type de neurotransmetteur). Grâce à ces récepteurs
qui s'ouvrent à l'arrivée des neurotransmetteurs, ceux-ci parviennent à pénétrer dans les dendrites du neurone récepteur de
l'influx. Et ainsi, le message nerveux peut passer d'un neurone à un autre.
| Constitution d'une synapse |
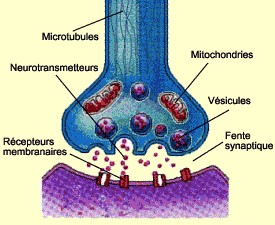
|
| (source: "le cerveau à tous les niveaux") |
Effet du neurotransmetteur
sur les récepteur membranaires :
les neurotransmetteurs, en se fixant
sur les récepteurs membranaires, peuvent avoir deux effets opposés sur ceux-ci. Soit qu'ils favorisent le passage de l'influx
nerveux entre les deux neurones, ce sont alors des neurones excitateurs, soit qu'ils diminuent la probabilité que l'influx
passe d'un neurone à un autre, on dit alors que les neurotransmetteurs sont inhibiteurs.
Prenons par exemple le cas du réflexe
myotatique.
| Le réflexe myotatique |
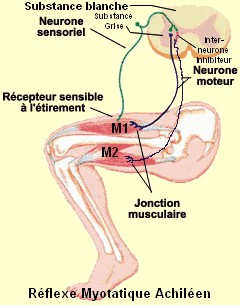
|
| (source: inspiré de "le cerveau à tous les niveaux" et modifié par nous-même) |
Lors du réflexe myotatique (contraction
d'un muscle en réponse à son propre étirement), le stimuli est l'étirement involontaire de M1. Ce stimuli est perçu au niveau
du récepteur, situé au niveau de M1. Un message circule alors à travers le neurone sensoriel (qui perçoit les stimuli provenant
du milieu environnant). Au niveau de la substance grise, le neurone sensoriel se divise en deux branches de neurone. La première
branche est directement mise en contact avec le neurone moteur (qui conduit l'influx nerveux au muscle, ce qui provoque son
mouvement ou son relâchement) de M1. Il y a contraction de M1, donc on peut en conclure que l'influx nerveux est passé du
neurone sensitif au neurone moteur de M1, ce qui signifie que les récepteurs membranaires du neurone moteur de M1 sont de
type excitateur. Par contre, la deuxième branche du neurone sensitif n'est pas directement mise en contact avec le neurone
moteur de M2, il y a entre les deux neurones une sorte de troisième neurone appelé interneurone. On remarque qu'il y a relâchement
de M2, ce qui rend compte d'une absence d'influx nerveux, ce qui voudrait dire que le message nerveux n'est pas passé du neurone
sensitif au neurone moteur de M2. On pourrait donc supposer que l'interneurone joue un rôle d'inhibiteur, et que donc ses
récepteurs membranaires sont de type inhibiteur.
II-2-3/ Le
sens de propagation du message nerveux :
Soit un neurone, que l'on nommera
neurone1, un neurone pré-synaptique (d'où vient l'influx nerveux), et soit un autre neurone, que l'on nommera neurone2, un
neurone post-synaptique (qui reçoit l'influx nerveux). Certains scientifiques se sont demandé si le neurone1 ne pouvait pas
recevoir l'influx nerveux du neurone2. Pour vérifier cela, ils ont procédé à plusieurs expériences que voilà.
Ils ont placé des électrodes stimulatrices
au niveau du corps cellulaire du neurone1, et des électrodes d'enregistrement au niveau de l'axone du neurone2, puis ils ont
regardé les enregistrement sur l'ossilloscope.
| Stimulation du neurone1 |
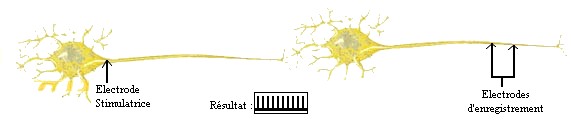
|
| (source: schéma réalisé par nous-même) |
On remarque qu'il y a une activité
enregistrée au niveau de l'axone du neurone2. Ceci veut dire qu'il y a changement de polarité, donc passage d'un influx nerveux.
Donc on peut en conclure que, comme on le sait déjà, l'influx nerveux passe du neurone1 vers le neurone2. Mais est-ce que
le contraire est possible?
Dans une deuxième expérience, ces
mêmes scientifiques inversent l'ordre de placement des électrodes. C'est-à-dire qu'ils placent les électrodes stimulatrices
au niveau du corps cellulaire du neurone2 et celles d'enregistrement au niveau de l'axone du neurone1. Ils relèvent l'enregistrement
de l'ossilloscope.
| Stimulation du neurone2 |
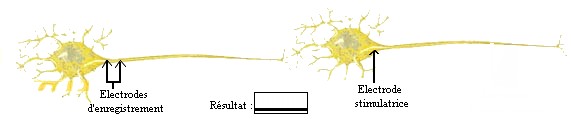
|
| (source: schéma réalisé par nous-même) |
On remarque qu'il n'y a aucune activité enregistrée au niveau de l'axone du neurone1. Ceci témoigne
d'une polarisation de l'axone, donc de la présence d'un potentiel de repos. Ce qui prouve que l'influx nerveux qui a pris
naissance au niveau du neurone2 n'est pas passé au neurone1. On peut donc en conclure que le sens de propagation de l'influx
nerveux se fait dans un seul sens, du neurone1 vers le neurone2.
|
|
|
|